

Optimize your Cellular Energy by Depleting Deuterium
Reduce Deuterium Interference by Drinking Litewater Daily
Reduce Deuterium Interference by Drinking Litewater Daily
Reduce Deuterium Interference by Drinking Litewater Daily
Reduce Deuterium Interference by Drinking Litewater Daily
Reduce Deuterium Interference by Drinking Litewater Daily
By leveling down deuterium with Litewater, the First Super Deuterium Depleted Water in the World. Available in 5 & 10 ppm (parts-per-million) total deuterium content to reduce your total body burden of deuterium at your own pace.
What is Litewater?
Litewater is the most deuterium depleted water (DDW) that exists. A lower concentration of deuterium compared to all other drinking water makes it the purest water in the world!
3-Month Mito Reset
Get a complete Litewater starter kit to Deuterium Depletion and to reset your mitochondria.
Benefits
Drinking Litewater deuterium depleted water allows you to lower your body's deuterium levels at your own pace and maintain the easiest most powerful anti-aging strategy that exists.
Science
From the first published study on deuterium depleted water in 1961 to the 2007 discovery of just how deuterium damages mitochondria, the geroprotective benefits of being deuterium depleted are clear.
Recommended Use
To deplete deuterium it is generally recognized that 45-90 days on Litewater is needed. Watch this 1 minute video. Answer these questions and find the strategy right for you. Click Here.
A Brief History of Deuterium Depleted Water
The story of the greatest discovery in biology and breakthrough for longevity and human health.
Test Your Levels ~ Deuterium Saliva Test
Find out your baseline of deuterium, then test after 2-3 months of depletion to check results. Know your level of this key biomarker of health and aging.
Frequently Asked Questions
Watch this Quick Explainer Video First. Deuterium is one of the three forms (called isotopes) of the first element, hydrogen. The first hydrogen isotope is named ‘protium’ (the most abundant by far and the lightest), consisting of a proton and an electron. Protium is commonly expressed by the symbol '1H’. The second isotope is named ‘deuterium’ (much less abundant than protium but twice as heavy due to the addition of a neutron). Deuterium is commonly expressed by the symbol ‘D’ or ‘ 2H’. The most important fact to know is that in nature, both protium and deuterium combine with oxygen to form water molecules.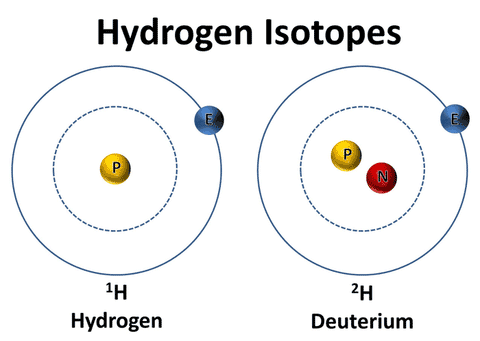
PREVALENCE OF HYDROGEN ISOTOPES ON EARTH:
Protium - 99.98% (by number)
Deuterium - 0.0156% (by number)
On our planet, for every 1,000,000 atoms of Hydrogen there are about 150 atoms of deuterium, (150 ppm)*.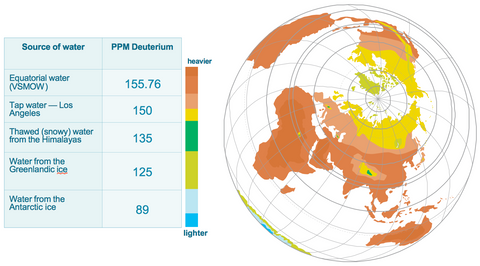
*‘Parts-per-million’ means ‘out of a million’. It is commonly used to describe the concentration of something in a solution of water. One ppm is equivalent to 1 milligram of something per liter of water.
What we commonly refer to as water, consume every day, use to prepare foods and beverages, irrigate our lawns, wash our clothes and put out fires, is more complex in its composition than is indicated by the popular chemical formula “H2O”. Chemically speaking, natural water predominantly consists of three molecules: H2O, HDO and D2O, and if you consider all the isotopes of oxygen and the various configurations, there are really 63 different types of water molecules known, in approximately the following ratios.
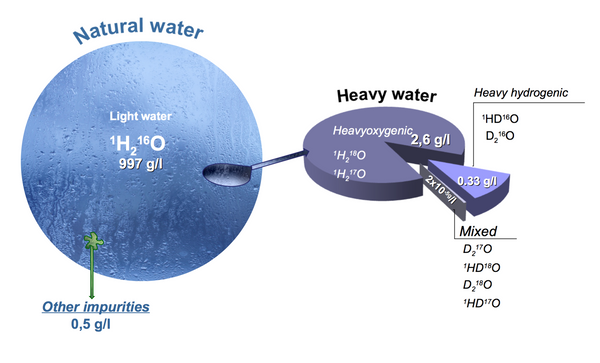
Deuterium depleted water (DDW), also referred to as light water is a term given to a unique composition of water in which the natural deuterium content is at least 15% less than 155 ppm, which the the deuterium content of the ocean and the highest level present in the natural waters on Earth. In a 1 Liter bottle of regular water there is about 6 drops of heavy water a.k.a, HDO.
Scientists, studying the cause of aging, discovered that even when a small amount of Deuterium containing Heavy Water is removed from normal water, the benefits are profound. Excess deuterium interferes in all biological process and is known to damage the mitochondria over time. Drinking deuterium depleted water allows you to reduce your total body burden of deuterium.
A few drops of heavy water in every glass of water we drink doesn't seem like much, but the cumulative effect is eye-opening, we have four times more deuterium in our blood than the most essential nutrients and minerals we require to live while deuterium slows down all reactions upto 9 times (9x)! This is known as the Kinetic Isotope Effect.
In 2007 it was published for the first time exactly how deuterium damages our ATP Synthase Nanomotors, recognized most beautiful nanomachine in nature, and the one responsible for producing ATP, (Adenosine-triphosphate) the molecule that produces the energy we need to live and breathe.
Litewater is our brand of deuterium-depleted water (DDW). It is water which has a lower concentration of deuterium than occurs naturally. Daily consumption of Litewater eliminates deuterium interference. Litewater has a 94-97% lower concentration of deuterium than regular water. Litewater is the most deuterium depleted water in the world which makes it ideal to be dilute with regular water from 1 to 4 times to obtain between 80 ppm -122 ppm deuterium depleted water. More value, less carbon footprint.
See Litewater Dilution Chart.
There is no ‘perfect” 100% deuterium depleted water with 0 ppm deuterium in nature. All natural water contains different concentrations of deuterium. The’ lightest’ water on Earth, formed as a result of natural meteorological processes, is the snow and ice in Antarctica, which is approximately 89 ppm, the closest to the water inside our cells which is 40-60 ppm.
Unfortunately, as perfect as this water would be to drink, its remote inaccessibility keeps it from being used as a water resource. Deuterium depleted water may only be obtained in large quantities by artificial means. Litewater Scientific processed ‘light water’ water far surpasses Antarctic water in purity.
Yes, deuterium depleted water can be used like any other drinking water for daily use with no age restrictions. Nevertheless, always seek the advice of your physician or qualified healthcare provider with any questions you may have regarding the introduction of anything new to your diet.


Litewater is a leader in DDW products. The quality and packaging are excellent, and the subscription service makes it easy to always have Litewater on hand.
~ Brian S.
Ever since the birth of our two kids (3 years and 3 months) my wife has struggled with not having energy to do basic household tasks. We've noticed a significant boost in her energy level since she started drinking Litewater two months ago. I'm very eager to share my enthusiasm about this product and deuterium depleted water in general. A secret that unlocks health like this is too good to keep to ourselves!
~ Matthew D.
The company makes awesome products. I really like having the option to purchase in glass bottles. Customer service is great too. The partnership with Hydroshot is nice too since DDW is used to make it.
~ Michael S.
I have struggled with food intolerances for years and upon using Litewater, I was able to add more foods into my diet. I'm such a big fan of deuterium depletion!
~ Miranda H.
I’ve been using Litewater for about a half year now. I’m going to keep using it. It's cheaper than going to the doctor continuously and getting no results. At 66 I’m feeling much healthier than my past 8 years. I’ve got better energy with much less aches and pains.
~ David W.
Litewater is essential for me in healing and maintaining my body and for improving my immune system. I have seen my labs and overall energy improve significantly since using it. As a Health Coach, I help others incorporate Litwater into their protocols and approach, especailly when they are experiencing challenges with dehydration.
~ Meghan O.
There is no doubt in my mind that this water works incredibly well. I higly recommend Litewater!
~ Gregory C.
Litewater is a leader in DDW products. The quality and packaging are excellent, and the subscription service makes it easy to always have Litewater on hand.
~ Brian S.
Ever since the birth of our two kids (3 years and 3 months) my wife has struggled with not having energy to do basic household tasks. We've noticed a significant boost in her energy level since she started drinking Litewater two months ago. I'm very eager to share my enthusiasm about this product and deuterium depleted water in general. A secret that unlocks health like this is too good to keep to ourselves!
~ Matthew D.
The company makes awesome products. I really like having the option to purchase in glass bottles. Customer service is great too. The partnership with Hydroshot is nice too since DDW is used to make it.
~ Michael S.
I have struggled with food intolerances for years and upon using Litewater, I was able to add more foods into my diet. I'm such a big fan of deuterium depletion!
~ Miranda H.
I’ve been using Litewater for about a half year now. I’m going to keep using it. It's cheaper than going to the doctor continuously and getting no results. At 66 I’m feeling much healthier than my past 8 years. I’ve got better energy with much less aches and pains.
~ David W.
Litewater is essential for me in healing and maintaining my body and for improving my immune system. I have seen my labs and overall energy improve significantly since using it. As a Health Coach, I help others incorporate Litwater into their protocols and approach, especailly when they are experiencing challenges with dehydration.
~ Meghan O.
There is no doubt in my mind that this water works incredibly well. I higly recommend Litewater!
~ Gregory C.
Litewater is a leader in DDW products. The quality and packaging are excellent, and the subscription service makes it easy to always have Litewater on hand.
~ Brian S.
Join our Deuterium Depleted Lifestyle Community!
